Influenza: The Master of Rapid Evolution
Preface
Influenza virus is a household name worldwide. Records of suspected influenza outbreaks date back to 500 BC. In modern history, the devastating 1918 influenza pandemic went down in history, infecting around one billion people and killing nearly 50 million. Another global pandemic, the Asian flu of 1957, the second major worldwide influenza outbreak of the 20th century, claimed roughly two million lives. Such catastrophic pandemics are closely linked to the virus’s wide range of host species.
Apart from the well-known avian and swine influenza, canine influenza is gradually gaining prevalence. Given dogs’ close companionship with humans, this highly contagious virus deserves adequate attention.
Etiology
Influenza Classification: Influenza viruses are categorized into four types: Influenza A, B, C and D. Influenza A is the most prevalent type, so the following discussion focuses primarily on Influenza A.
Influenza viruses belong to the Orthomyxoviridae family and are single-stranded RNA viruses. Subtypes of Influenza A are defined by the antigenic properties of two surface glycoproteins: hemagglutinin (H1–H18) and neuraminidase (N1–N11).
The influenza viral genome consists of eight discrete gene segments that enable extensive genetic reassortment of one or more genomic fragments. Co-infection of a single host with influenza viruses from different species facilitates viral gene reassortment, generating reassortant strains capable of adapting to new hosts and even crossing species barriers. For instance, the influenza pandemics in 1957 and 1968 stemmed from genetic reassortment between avian and human influenza viruses.
Epidemiology
Influenza viruses have an extensive host range, predominantly infecting mammals including humans, horses, dogs and swine, as well as avian species such as wild waterfowl and domestic poultry.
Canine influenza viruses (CIV) have undergone rapid evolutionary changes. Notably, CIV originating via host jump mutation exhibits greater mutability than its progenitor strains. As companion animals closely associated with humans, dogs pose public health concerns due to the high genetic variability of CIV.
1.H3N8 Subtype:
Evidence dating back to 1999 confirmed interspecies transmission of equine-origin H3N8 to dogs. A large outbreak occurred in racing greyhounds in Florida in 2004, with 22 infected and 8 fatal cases. Subsequent serological screening of clinic-owned dogs across Florida found 97% of tested animals seropositive against the identical virus, verifying sustained dog-to-dog horizontal transmission. Isolates of the Florida H3N8 strain were verified as host-adapted canine influenza virus. Reports of H3N8 infections have declined progressively since 2016.
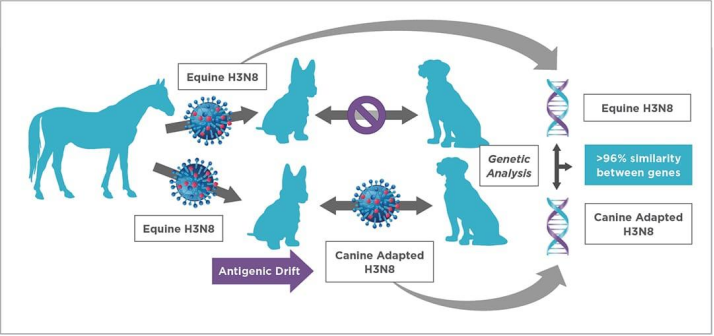
2.H3N2 Subtype:
Canine influenza virus H3N2 was first identified in China and South Korea in 2006 and confirmed to be of avian origin. The H3N2 influenza virus was introduced from Asia to the Chicago region of the United States in 2015, triggering extensive disease outbreaks among dogs thereafter. Despite local control interventions, the virus persisted in canine populations in and around Chicago before spreading to southeastern states including Georgia and North Carolina. Successive outbreaks kept emerging afterwards, prompting the United States to tighten entry regulations for imported dogs.
A US research found that following the emergence of canine H3N2 influenza virus (CIV), the virus has accumulated more non-synonymous substitutions at all sites compared with its ancestral avian H3N2 influenza virus (AIV). The average dN/dS ratio (ratio of non-synonymous to synonymous evolutionary changes) across major coding regions ranges from 0.15 to 0.26 for CIV, versus 0.03 to 0.10 for avian-origin H3N2 AIV. These elevated average dN/dS values in CIV correspond to abundant amino acid alterations at the protein level accumulated during its circulation in Asia and even within its relatively short transmission period across the United States.
3.H3N6 Subtype:
During 2018–2019, an active epidemiological surveillance study on canine influenza virus (CIV) in Liaoning Province identified and confirmed a novel CIV subtype H3N6. Genetic and phylogenetic analyses revealed that the NA gene of H3N6 CIV clusters with that of avian H5N6 influenza virus (AIV). Researchers hypothesize that this novel H3N6 CIV most likely originated from genetic reassortment between H3N2 CIV and H5N6 AIV, and interspecies reassortment events of H5N6 AIV have been documented previously.
The study also demonstrated that CIV evolves and adapts to new hosts at a remarkably rapid pace. H3N2 CIV isolated in November 2018 displayed restricted replicative capacity both in vitro and in vivo. However, isolates collected only three months later (March 2019) exhibited markedly enhanced replication efficiency. Furthermore, the reassortant H3N6 CIV emerged in April 2019, just one month afterward, featuring more stable host replication compared with H3N2 CIV.

- X-axis: Viral load
- Y-axis: Collected tissue sites
Transmission
Like other influenza A viruses, canine influenza virus (CIV) spreads mainly via direct and indirect contact with respiratory secretions and infectious aerosols. The incubation period lasts 2 to 3 days with high levels of viral shedding throughout this phase. Viral excretion drops rapidly once clinical signs develop after incubation. In one cohort of shelter dogs infected in Chicago, H3N2 CIV was detected in nasal secretions for more than 21 days post-infection. Notably, infected dogs shed virus before the onset of clinical symptoms and may continue shedding after clinical resolution; for H3N2 subtype, shedding can persist for over three weeks. Group housing is a major predisposing risk factor for viral transmission.
Clinical Signs
Symptoms caused by CIV are similar to canine infectious respiratory disease. Infected dogs mainly develop upper respiratory manifestations including sneezing and nasal discharge (serous or mucopurulent), alongside systemic symptoms such as fever, inappetence and lethargy.
Diagnosis
PCR is the preferred clinical assay for acute infection, with existing tests capable of detecting both H3N8 and H3N2 CIV subtypes. Sampling is recommended within the first 4–5 days after symptom onset, corresponding to peak viral shedding. Specimens collected at late disease stages carry an elevated risk of false-negative results due to diminished viral excretion; however, prolonged shedding of H3N2 enables reliable testing even at later phases of illness. Nasopharyngeal swabs are the preferred sample type.
In cases of false-negative PCR findings, serological testing is recommended as an adjunct diagnostic method. Diagnosis is confirmed by a ≥4-fold rise in CIV antibody titer between acute-phase and convalescent-phase paired sera.
References
- Greene’s Infectious Diseases of the Dog and Cat; Chapter 25.
- Anderson TC, Crawford PC, Dubovi EJ, et al. Serological evidence of H3N8 canine influenza-like virus circulation in USA dogs prior to 2004. Vet J. 2012;191(3):312-316.
- Crawford PC, Dubovi EJ, Castleman WL, et al. Transmission of equine influenza virus to dogs. Science. 2005;310(5747):482-485.
- Meng B, Li H, Feng C, Guo W, Feng Y, Zhu D, Chen H, Zhang Y. Emergence of a novel reassortant H3N6 canine influenza virus. Frontiers in Microbiology. 2023;14.